

400-820-8531




N6-甲基腺苷(m6A)是真核生物mRNA中最重要的转录后修饰,在真核生物的病理和生理过程中起着至关重要的作用。糖尿病认知障碍是一种不可逆的中枢神经系统退行性疾病,目前发展的确切机制尚未完全阐明,并且缺乏有效的预防和治疗手段。
2024年2月6日,北京大学医学部联合首都医科大学附属北京朝阳医院在International Journal of Molecular Sciences上发表题为“Altered N6-Methyladenosine modicification patterns and transcript profiles contributes to cognitive dysfunction in high-fat induced diabetic mice”的研究文章。研究人员探讨了高脂肪饮食对葡萄糖代谢的干扰会导致与m6A修饰相关的酶的表达发生变化的相关机制。该成果中拜谱生物为其提供m6A甲基化分析技术服务,为糖尿病小鼠高脂肪饮食诱导的认知功能障碍机制提供了新的线索。

文章名称:Altered N6-Methyladenosine modicification patterns and transcript profiles contributes to cognitive dysfunction in high-fat induced diabetic mice(Int J Mol Sci.,IF=6.2,2024.2)
客户单位:北京大学医学部、首都医科大学附属北京朝阳医院
研究材料:小鼠脑组织
拜谱提供技术:m6A甲基化(MeRIP-seq)
摘要图:
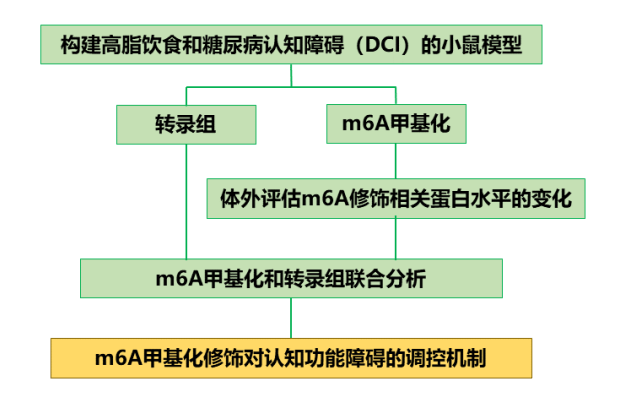 图1| 摘要图(图源:Zhaoming Cao, et al., Int J Mol Sci., 2024)
图1| 摘要图(图源:Zhaoming Cao, et al., Int J Mol Sci., 2024)
研究结果
1.DCI小鼠海马中m6A基因修饰的改变
研究人员首先建立了高脂饮食和认知障碍(DCI)的小鼠模型。为比较对照组(n = 3)和DCI组(n = 3)之间的m6A基因修饰,研究人员使用MeRIP-seq进行了转录组范围内的m6A-seq分析。在DCI组的10664个编码基因转录物(mRNAs)中共发现对照组的23711个不重叠的m6A峰。相比之下,DCI组表现出23,751个非重叠m6A峰。其中,9345个甲基化基因在对照组和DCI组之间重叠(图2)。
 图2| 对照组和DCl组m6a修饰基因的维恩图
图2| 对照组和DCl组m6a修饰基因的维恩图
共鉴定出1278个差异甲基化的m6A位点(DMMSs),其中60.88%(778/1278)和39.12%(500/1278)的甲基化水平分别显著高于对照组。分析其分布情况发现m6A峰分布在整个RNA中(图3A),并根据其分布的转录区域分为以下五组:5'UTR、3'UTR、转录起始位点区域片段、终止密码子片段、起始密码子片段和编码序列(CDS)。m6A峰的在3'UTR、CDS和终止密码子附近显著富集(图3B)。为了评估这些基因上m6A修饰的区域,研究人员筛选了有峰注释的区域,并测定了这些基因上reads的覆盖率。在DCI组中,m6A修饰区偏向于3'UTR区(图3C)。所有mRNA内的DMMS都被定位到染色体上去评估其分布概况。不同小鼠染色体的m6A修饰谱不同。
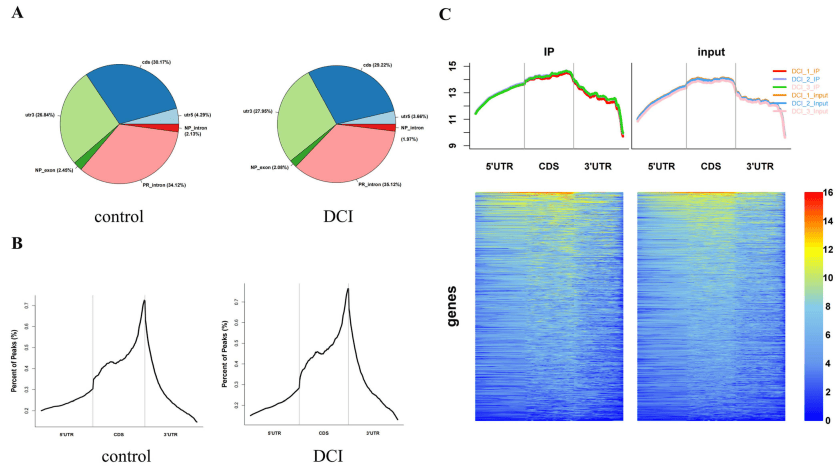 图3| 小鼠海马中m6A甲基化图谱的改变
图3| 小鼠海马中m6A甲基化图谱的改变
为了检验m6A修饰在DCI小鼠中的生物学功能,研究人员从两组中选择含有DMMS的蛋白质编码基因进行GO和KEGG分析(图4A)。在GO分析中,高甲基化基因在细胞过程、生物过程调控、催化活性和杂环化合物结合中显著富集。KEGG通路分析显示,高甲基化基因与T细胞受体信号通路、小细胞肺癌和RIG-I样受体信号通路相关(图4B)。对于低甲基化基因,GO分析显示在细胞过程、代谢过程和结合过程中显著富集有机环化合物、结合杂环化合物和催化活性(图4C)。KEGG通路分析揭示了低甲基化基因与肥厚性心肌病和Hippo信号通路的关联(图4D)。
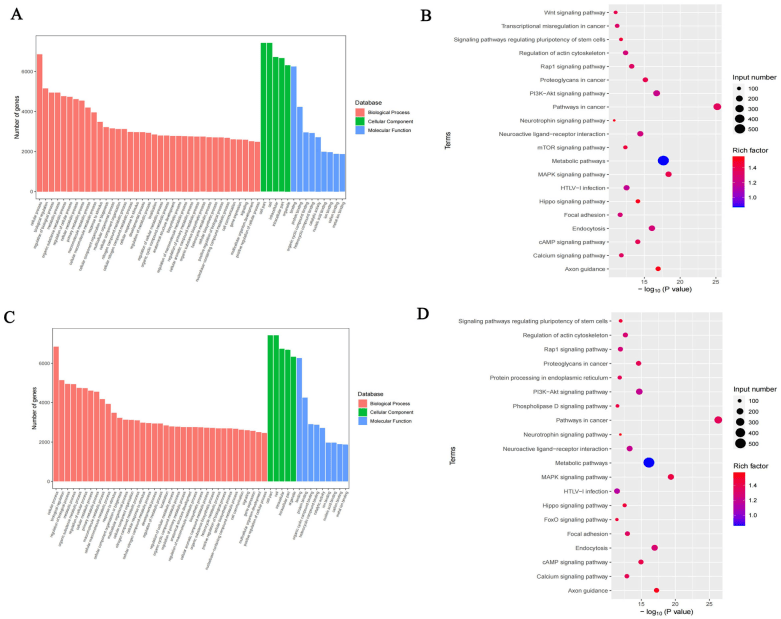 图4| 差异甲基化m6A位点的通路富集
图4| 差异甲基化m6A位点的通路富集
2.m6A MeRIP-Seq和RNA-Seq数据的联合分析
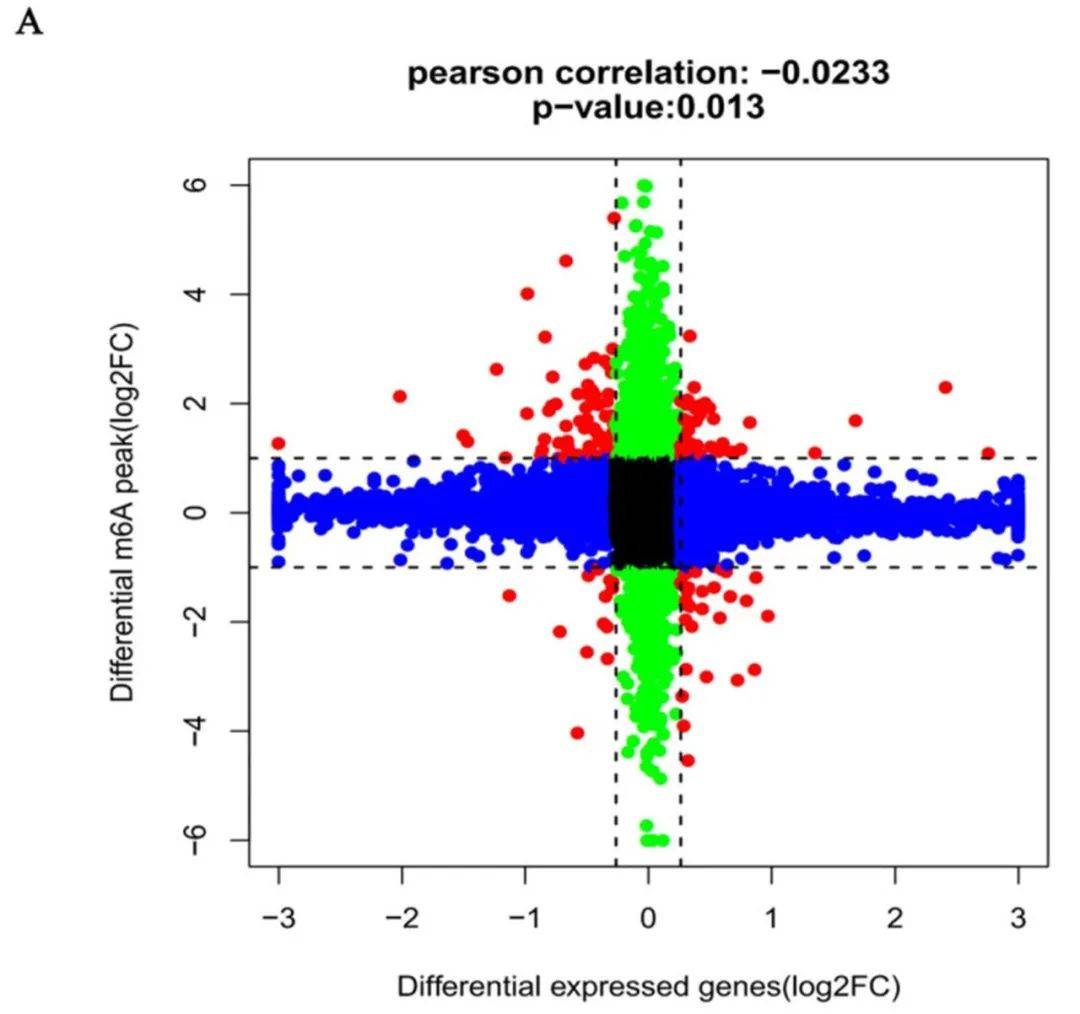
随后研究人员将m6A MeRIP-Seq的数据和先前RNA-Seq的数据进行联合分析,发现163个mRNA,m6A峰和mRNA水平显著改变。41种mRNA的m6A和mRNA水平上调,14种mRNA水平下调。此外,30个基因mRNA表达上调,m6A水平下调,78个基因mRNA表达下调,m6A水平上调(图5A)。最后,构建了一个PPI网络来显示163个基因编码的蛋白之间的连接(图5B)。

如图6所示,GO和KEGG分析分为四组:(1) m6A上调,mRNA下调;(2) m6A上调,mRNA上调;(3) m6A下调,mRNA下调;(4) 基因m6A下调和mRNA上调。在GO分析中,m6A上调和mRNA下调的基因在细胞代谢过程、芳香族化合物生物合成过程和RNA代谢过程的调控中显著富集。KEGG分析显示,这些基因在脂肪酸代谢、小细胞肺癌、丙酸代谢和丙酮酸代谢中富集。对于m6A上调和mRNA上调的基因,GO分析显示在亚铁结合、亚胺丙氨酸脱羧酶活性、天冬氨酸1-脱羧酶活性和细胞连接组装负调控中富集;KEGG分析显示在2型糖尿病、脂肪细胞因子信号通路和胰岛素信号通路中富集。在GO分析中,m6A下调和mRNA下调的基因在寡肽结合、谷胱甘肽结合和二硫氧化物氧化还原酶活性中富集,而在KEGG分析中,它们在柠檬酸循环(TCA循环)、丙酸代谢和多物种凋亡中富集。在GO分析中,m6A下调和mRNA上调的基因在胚胎骨关节形态发生、蛋白激酶活性激活和NADP代谢过程中富集,在KEGG分析中,m6A下调和mRNA上调基因在新霉素、卡那霉素和神经营养因子信号通路中富集。
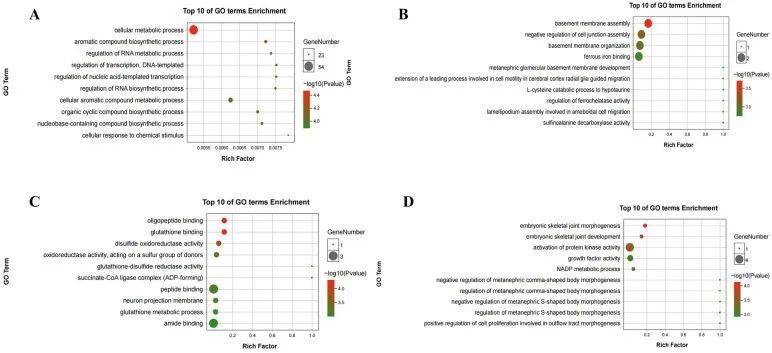 图5| m6A甲基化与mRNA表达的联合分析
图5| m6A甲基化与mRNA表达的联合分析
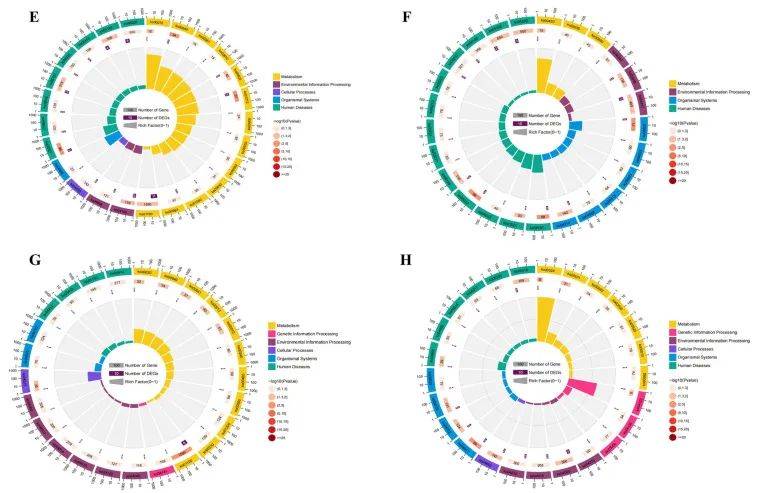 图6| 163个DEGs和DMMS的GO和KEGG分析
图6| 163个DEGs和DMMS的GO和KEGG分析
拜谱生物小结
本研究表明高脂饮食对葡萄糖代谢的干扰会导致与m6A修饰相关的酶的表达发生变化。这些改变导致与神经退行性疾病相关的基因甲基化模式的改变,特别是在小鼠的海马区。这些发现为高脂肪饮食诱导糖尿病小鼠认知功能障碍的机制提供了新的见解,可能为糖尿病诱导的海马病变提供新的治疗策略。这一过程中拜谱生物提供了m6A甲基化技术服务,拜谱生物可提供完善成熟的蛋白组学、代谢组学、转录组学以及多组学联合产品技术服务,欢迎致电咨询!
参考文献:Cao Z, An Y, Lu Y. Altered N6-Methyladenosine Modification Patterns and Transcript Profiles Contributes to Cognitive Dysfunction in High-Fat Induced Diabetic Mice. Int J Mol Sci. 2024. doi: 10.3390/ijms25041990.





关注公众号





关注小红书


