

400-820-8531




背景介绍
泛素信号通路长期被认为主要修饰蛋白质,但糖类、脂质、核苷酸等非蛋白类物质的泛素化长期被忽视。传统泛素组学与蛋白质组学技术无法识别非蛋白底物的泛素修饰,导致这类修饰的分布、丰度与功能始终不明确。
2026年4月,《Nature》发表一项突破性研究,建立NoPro‑clipping(非蛋白质泛素剪切)质谱新方法,首次在哺乳动物细胞与组织中系统性证实非蛋白物质的泛素化广泛存在。研究发现,糖原在所有含糖原组织中均被泛素修饰,且泛素化可介导糖原进入溶酶体降解;肝脏在禁食状态下,糖原消耗与糖原泛素化上升同步发生。此外,研究还鉴定出甘油、精胺两种新型泛素化底物,正式将泛素从“蛋白质修饰因子”提升为全生物分子通用修饰因子。

NoPro-clipping技术
建立非蛋白质泛素化检测方法
研究团队开发了NoPro‑clipping技术,可以高灵敏地检测非蛋白底物上的泛素化。该方法将泛素剪切酶与分选酶标记结合,先通过尺寸排阻柱去除<7kDa小分子,再用Lbpro* 或BpJOS剪切酶处理,使泛素在底物上保留C端GG标签;第二次过滤(<3 kDa)去除蛋白质等大分子,富集GG标记的小分子底物后进行质谱检测。为提升灵敏度,研究引入分选酶 A介导的ClipTag标记(生物素标记的LPXTG肽段),将GG修饰底物转化为肽样结构,实现固相富集与纳流质谱分析。团队利用HOIL‑1L体外合成泛素化麦芽糖、麦芽七糖标准品,验证流程可靠性,并通过α-淀粉酶将大分子泛素化糖原降解为GG修饰的麦芽三糖和麦芽四糖,使其适配质谱检测。体外标准品与细胞裂解物掺入实验均验证了方法在复杂体系中的回收率与特异性。
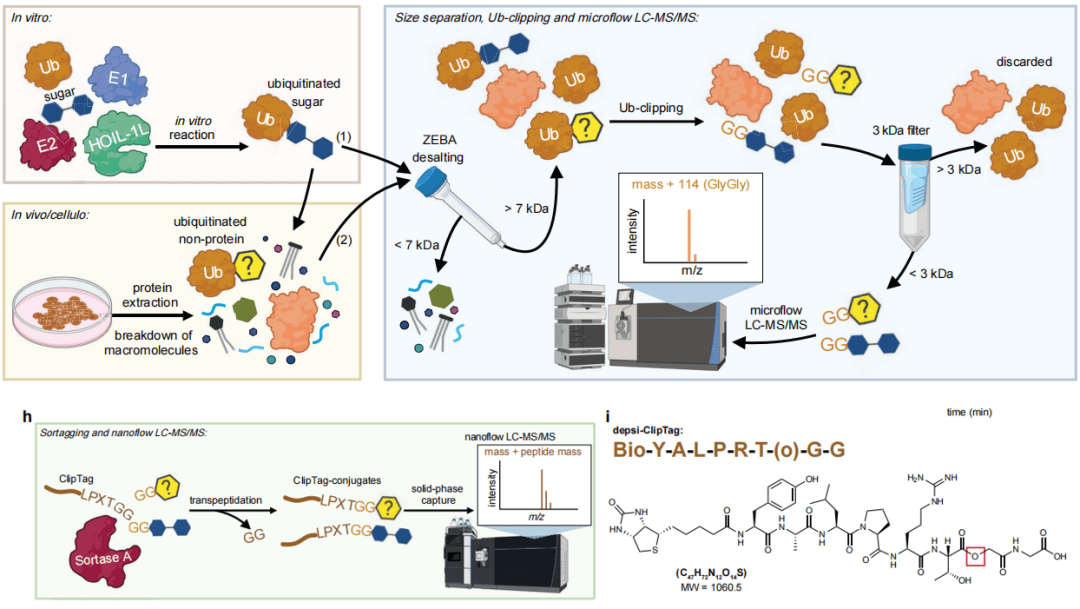
图1 NoPro-clipping工作流程与糖原泛素化检测
诱导糖原泛素化
细胞中糖原泛素化的验证
为在细胞内证实糖原泛素化,研究构建FKBP/FRB化学邻近诱导系统。在人肝癌Huh7细胞中共表达FKBP‑GFP‑CBM20与mCherry‑FRB‑HOIL‑1L,加入雷帕霉素后,GFP 标记的糖原颗粒形成荧光焦点,并与HOIL‑1L、泛素信号共定位。该信号依赖两种质粒共表达与完整泛素系统,E1抑制剂TAK243可完全阻断。经NoPro‑clipping检测,细胞内产生与体外泛素化糖原一致的ClipTag‑葡寡糖信号,MS2碎片图谱高度匹配。进一步通过¹³C₆-葡萄糖代谢标记,结合自激活HOIL-1L*过表达,研究者在细胞内检测到¹³C标记的泛素化糖原,质谱显示明确的质量位移,为细胞内源性糖原泛素化提供了独立证据。
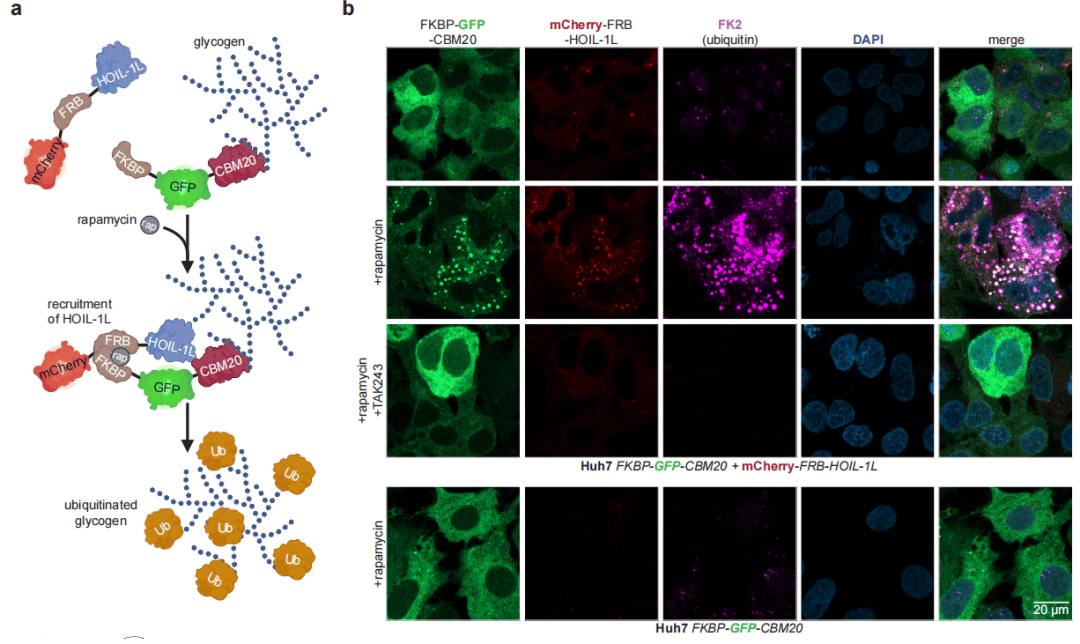
图2 邻近诱导糖原泛素化的细胞验证
Ub-糖原触发溶酶体转运
泛素依赖性糖原清除新途径
糖原的溶酶体降解早已被发现,但泛素化的调控作用此前未被揭示。为排除雷帕霉素对自噬的干扰,研究改用PYR/ABI-化学诱导剂系统(使用mandipropamid作为二聚化诱导剂)。在Huh7细胞中,诱导后糖原、HOIL‑1L与泛素共定位,并与溶酶体标记LAMP1共定位,而不与内体标记EEA1共定位;巴弗洛霉素A1预处理可增强共定位信号,TAK243或催化失活突变HOIL‑1L H510A则完全阻断转运。在高糖原卵巢癌细胞SKOV3中,常规培养即可检测内源性泛素化糖原,巴弗洛霉素A1处理使糖原与泛素化糖原均上升约3倍;邻近诱导后糖原水平下降约25%。结果证明,泛素化是糖原靶向溶酶体降解的关键信号。
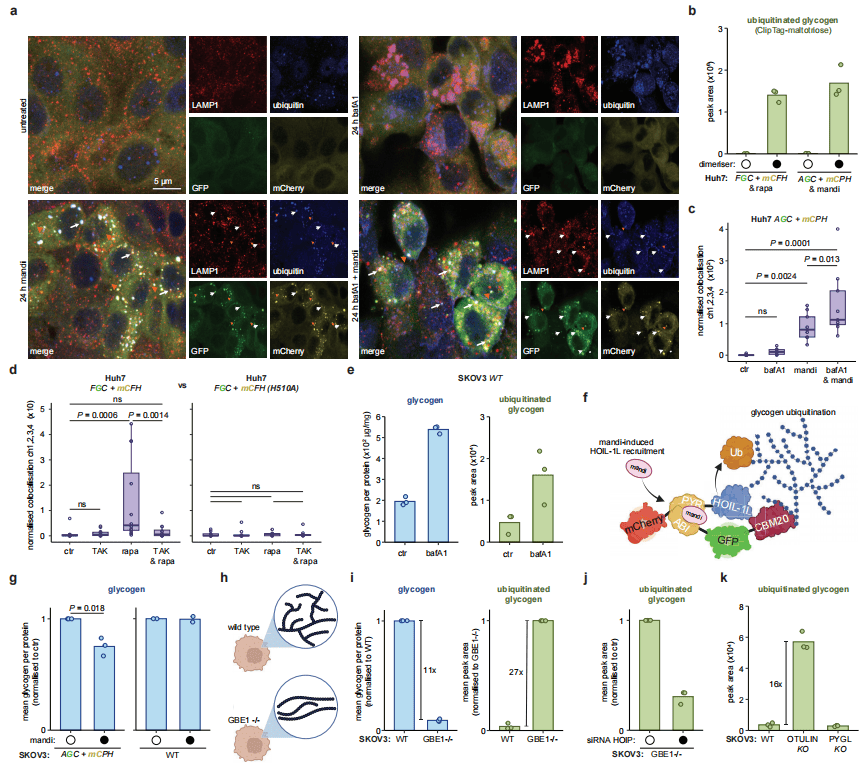
图3 泛素依赖性糖原溶酶体清除
糖原贮积病中的泛素化改变
LUBAC-OTULIN轴对异常糖原的调控
研究进一步探索糖原贮积病(GSD)中泛素化的作用。糖原分支酶GBE1负责维持糖原高度分支结构,在SKOV3细胞中敲除GBE1后,可检测糖原下降11倍,并出现GSD IV特征性多聚葡聚糖体(PSBs),而泛素化糖原显著上升27倍,归一化后上升近300倍,提示异常糖原被选择性泛素化标记。机制上,LUBAC复合物(含HOIL‑1L、HOIP)催化糖原泛素化,敲低HOIP可使泛素化糖原下降约60%;去泛素酶OTULIN特异性水解M1 泛素链,敲除OTULIN后泛素化糖原上升16倍,而糖原总量不变。OTULIN不能直接水解泛素‑葡萄糖键,说明其通过调控M1链间接控制糖原泛素化。
禁食反应中的泛素化糖原
生理条件下糖原泛素化的动态调控
为明确生理意义,研究在野生型C57BL/6小鼠多组织中检测泛素化糖原,结果显示脑、心脏、肺、肝脏、骨骼肌均存在修饰,其中肝脏与骨骼肌丰度最高。禁食实验显示,肝脏糖原在禁食6h下降约90%,24–48h接近耗尽;而泛素化糖原在6h禁食后显著上升,相对水平升高8倍,提示泛素化主动参与糖原分解。再喂食3h后糖原快速恢复,但泛素化糖原不回升,24h后才恢复至基础水平。定量显示,禁食6h时 **>1% 的总泛素结合在糖原上 **,含量达1.35pmol/mg 蛋白,与细胞内K11泛素链丰度相当;禁食24h后降至72fmol/mg。研究还在人骨骼肌活检样本中检测到泛素化糖原,具备临床转化潜力。
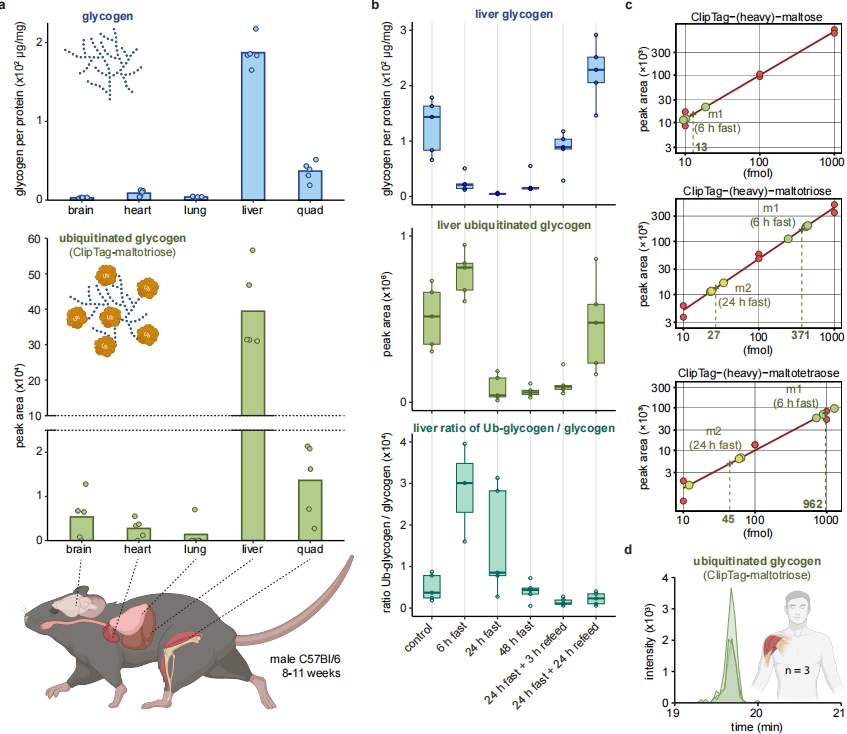
图4 小鼠组织中泛素化糖原的动态调控及人骨骼肌验证
其他代谢物的泛素化发现
非靶向NoPro-clipping发现泛素化甘油和精胺
在靶向分析糖原基础上,研究采用非靶向NoPro‑clipping拓展底物谱。在Huh7细胞中过表达HOIL‑1L*,对94027个MS1信号进行三重筛选:依赖BpJOS剪切、依赖ClipTag标记、为细胞裂解物特有,最终得到185个特征,其中包含已知ClipTag‑葡寡糖,验证方法可靠。其中两个信号ΔMW分别为92.0458与202.2159,匹配甘油与精胺。体外合成泛素化甘油、精胺标准品,其保留时间、m/z、MS2图谱与内源性信号完全一致;¹³C葡萄糖标记证实甘油泛素化来自细胞代谢,DFMO抑制精胺合成可降低泛素化精胺。结果支持甘油与精胺为内源性泛素化底物。
拜谱小结
本研究以NoPro‑clipping技术为核心,构建了 “方法建立—细胞验证—疾病机制—生理调控—新底物拓展” 的研究框架,首次系统性证明非蛋白生物分子广泛存在泛素化修饰。研究明确糖原泛素化由LUBAC‑OTULIN轴调控,介导溶酶体降解,参与禁食应答与糖原贮积病病理过程,并发现甘油、精胺两种新型小分子泛素化底物。该成果突破泛素仅修饰蛋白质的传统认知,将其定义为全生物分子通用修饰,为糖原贮积症、代谢综合征、脂肪肝等疾病提供全新机制解释与药物靶点。
参考文献
Jochem M, Cobbold SA, Goodman CA, et al. Ubiquitination of glycogen and metabolites in cells and tissues. Nature (2026). https://doi.org/10.1038/s41586-026-10548-x





关注公众号





关注小红书


