

400-820-8531




脂质是必不可少的细胞成分,在细胞生物学的多个方面发挥作用。理论上,脂质大约有10万种,其中细胞内存在数千种化学性质不同的脂质种类。脂质的各种生物物理特性使膜能够改变其形状、厚度和顺序。这些特性对于分裂、融合、出芽、成管和脂筏的形成是必不可少的。脂质还具有多种生化作用,包括参与翻译后修饰(如肉豆蔻化、棕榈酰化),作为能量代谢和产热的底物等。虽然在脂质结构多样性和确定特定脂质功能方面取得了重大进展,但目前对脂质多样性及细胞脂质的组成差异如何影响细胞生物学以及它如何有助于细胞特异性功能的了解有限。
2024年4月澳大利亚贝克心脏和糖尿病研究所的Andrew J. Murphy、Graeme I. Lancaster研究团队在Nature cell biology(17.3)杂志上发表题为“A lipid atlas of human and mouse immune cells provides insights into ferroptosis susceptibility”文章。该文章采用高通量靶向脂质组学技术证明了每种免疫细胞都具有其独特的脂质特征,还发现含多不饱和脂肪酸的甘油磷脂(PUFA-PL)含量的差异是免疫细胞对铁死亡不同易感性的基础,并且低PUFA-PL含量可以增强活化中性粒细胞对铁死亡的抵抗力。拜谱生物近期推出了MLT4500医学高通量靶向脂质组产品,多针检测4500+脂质分子,其中甘油磷脂类3100+,提供绝对定量数据和丰富的数据分析内容。敬请咨询!

英文标题:A lipid atlas of human and mouse immune cells provides insights into ferroptosis susceptibility
期刊:Nature cell biology
影响因子:17.3
发表时间:2024年4月
作者单位:澳大利亚贝克心脏和糖尿病研究所
研究材料:人外周血的16种成熟免疫细胞、小鼠外周血的8种免疫细胞
组学技术:靶向脂质组学
一、研究思路

图1 人类(上)和小鼠(下)免疫细胞脂质图谱分析
(图源:Morgan, Pooranee K et al., Nat Cell Biol. 2024)
二、研究结果
01.人类免疫系统的脂质组学细胞图谱
为了验证细胞脂质组是人类免疫细胞的一个决定性特征的假设,作者从14名健康供体的外周血中分离了16个不同的成熟免疫细胞群,代表了所有主要的免疫细胞谱系及其亚群。通过超高效液相色谱与串联质谱联用(UHPLC ESI-MS/MS)对其细胞脂质进行了表征。共定量了524种脂质,其中505种在至少两种细胞类型之间有显著差异,属于多个不同的脂质类别,包括二酰基、醚和乙烯基醚甘油磷脂(PLs)(磷脂酰胆碱-PC,磷脂酰丝氨酸-PS,磷脂酰乙醇胺-PE,磷脂酰肌醇-PI,磷脂酰甘油-PA)、鞘脂(SLs)(鞘磷脂-SM)、神经酰胺、神经节苷脂、己糖神经酰胺、溶血磷脂(Lyso-PLs)、胆固醇(COH)。在目前的研究中,作者对膜脂质特别感兴趣,因此没有分析中性脂质,例如三酰甘油(TG)、二酰甘油(DG)和胆固醇酯(CE ),它们是脂滴的丰富成分。通过t-分布随机邻近嵌入(t-SNE)对整体脂质组进行可视化,证明了整体细胞脂质组是人类免疫系统细胞的识别特征(图2b)。
在骨髓细胞中SL和COH的比例较高,特别是在嗜中性粒细胞和嗜酸性粒细胞中,PL的比例相应降低(图2c),PLs是细胞膜中存在的最丰富的脂质类别(图2c ),但免疫细胞类型之间PL类别的比例差异不显著(图2d)。嗜中性粒细胞中PS的比例增加。与NK细胞和骨髓细胞相比,T和B淋巴细胞中PG的比例明显更高(图2d)。结果表明人类免疫细胞具有独特的磷脂组成。
进一步研究发现骨髓细胞中PC(O)、PC(P)、PE(O)和PE(P)的比例显著高于淋巴细胞(图2e、f)。PE和PC中二酰基、烷基醚和烯基醚组成的这些差异是在免疫细胞类型之间观察到的整体脂质组差异的主要驱动因素。
PL多样性的另一个主要驱动因素是酰基/烷基链的组成,其长度通常从16到24个碳原子不等,包含0到6个双键。这些差异在功能上很重要,因为它们会影响膜的生物物理和生化特性1。饱和的16:0和18:0链占整个PL池的很大比例(~50%),这在免疫细胞类型之间相对一致(图2g)。综上所述,PL组成的差异是人类免疫细胞脂质组的一个决定性特征。具体来说,化学键的使用差异(即:酯、醚和乙烯基醚)和烷基/酰基链多样性是免疫细胞类型之间脂质组差异的主要驱动因素。并证明免疫细胞在激活或成熟时经历脂质重塑(图2h-l)。
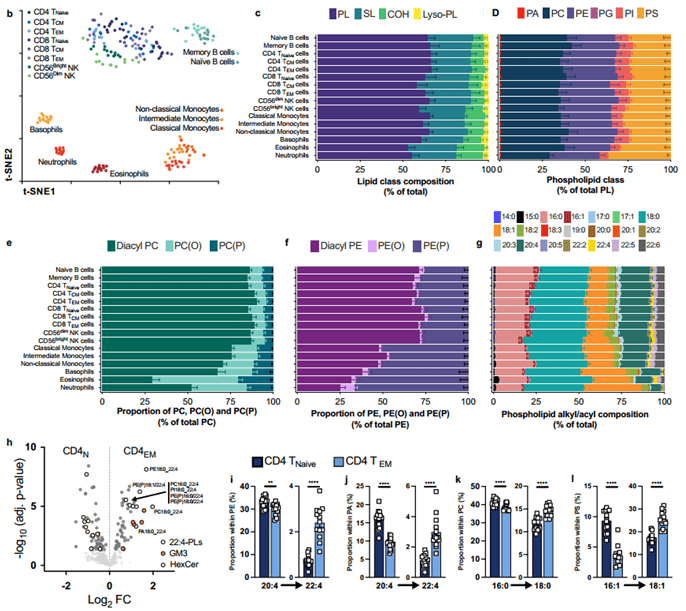
图2 人类免疫系统的脂质组学细胞图谱
(图源:Morgan, Pooranee K et al., Nat Cell Biol. 2024)
02.小鼠免疫系统的脂质组学细胞图谱
为了确定在人类免疫细胞中观察到的脂质组学特征是否存在于鼠免疫细胞中,作者构建了鼠免疫系统的脂质组学图谱。从8种不同的免疫细胞类型共检出524种脂质,400种在至少两种细胞类型之间有显著差异。使用t-SNE对不同鼠免疫细胞类型的整体脂质组进行可视化,揭示了髓样细胞和淋巴样细胞之间以及髓样细胞类型之间的明显区别(图3b)。
与人类免疫细胞相似,在PL、SL、lyso-PL和胆固醇的总比例或主要PL类别的比例中没有明显的细胞谱系或细胞类型差异(图3c,d)。PC和PE类中二酰基-PC/PE、醚PC(O)/PE(O)和乙烯基醚PC(P)/PE(P)的比例变化与在同等人类免疫细胞中观察到的变化一致(图3e、f)。因此,在细胞类型(主要是髓样与淋巴样)之间利用不同的sn1化学键(酯、醚、乙烯基醚键)是人类和小鼠免疫系统的保守特征。
接下来,作者研究了小鼠免疫细胞之间整体PL烷基/酰基组成的多样性。虽然在特定细胞类型之间存在许多显著差异,但没有观察到明显的谱系差异(图3g),但22:6除外,骨髓细胞的比例明显低于淋巴细胞。
对小鼠和人类免疫细胞脂质体之间的关系进行了皮尔逊相关系数检验。结果表明,人和小鼠免疫细胞的总体脂质丰度之间存在很强的相关性(中性粒细胞,r = 0.78;T 细胞,r = 0.80;B 细胞,r = 0.83)(图3h-j)。表明免疫细胞脂质组在小鼠和人类之间是保守的。

图3 小鼠免疫系统的脂质组学细胞图谱
(图源:Morgan, Pooranee K et al., Nat Cell Biol. 2024)
03.人类中性粒细胞和T细胞对铁死亡的敏感性不同
小鼠和人免疫细胞脂质组成的最显著特征之一是发现骨髓细胞富含ether-PLs(图2e,f和3e,f)。作者评估了在骨髓细胞发育过程中ether-PLs含量的变化。结果表明中性粒细胞发育过程中的转录重编程可能有助于脂质组成的改变。
免疫细胞脂质组成的最显著特征之一是髓样(低)和淋巴样(高)细胞之间PUFA比例的显著差异,特别是高度不饱和的20:4和22:6 PUFA。PUFA-PLs易受铁依赖性脂质过氧化的影响,这会导致一种被称为铁死亡的细胞死亡形式。结果发现PUFA-PLs 丰度的差异是免疫细胞易受脂质过氧化和铁死亡影响的主要决定因素。并且人类中性粒细胞和T细胞对铁死亡的敏感性不同。
04.改变细胞脂质组成会改变免疫细胞对脂质过氧化和铁死亡的易感性
数据表明,免疫细胞的PUFA含量决定了它们对铁死亡的易感性。作者假设改变内源性PUFA水平会改变铁死亡的易感性。首先,作者给T细胞补充单不饱和脂肪酸(MUFA)油酸(OA);18:1)或PL PE(18:0/18:1),以减少PUFA-PLs在膜中的比例。油酸酯和PE(18:0/18:1)都使T细胞PL组成向富含MUFA的方向转变,同时降低了含有4个碳碳双键的PLs的比例,例如含有20:4的PLs(图4a,b)。重要的是,补充OA和PE(18:0/18:1)都可以保护T细胞免受GPX4抑制引发的脂质过氧化和铁死亡(图4c,d)。接下来,作者在中性粒细胞中添加花生四烯酸(20:4)和二十二碳六烯酸(22:6)(AA + DHA)以增加内源性PUFA水平。AA + DHA处理降低了含1碳碳双键和2碳碳双键的PLs的比例,同时增加了含5、8和10碳碳双键的PLs的比例,例如20:4和22:6的PLs(图4e,f)。值得注意的是,在不使用ML210治疗的情况下,补充AA + DHA会导致剂量依赖性细胞死亡,并显著增加中性粒细胞的脂质过氧化(图4g,h)。这些作用在很大程度上被中性粒细胞与Fer-1共同处理而不是细胞凋亡或坏死性凋亡抑制剂逆转,表明真正的铁死亡(图4i,j)。总的来说,这些数据表明,PUFA丰度的差异导致了免疫细胞对脂质过氧化和铁死亡的不同敏感性。
NADPH氧化酶家族的酶(NOX1-5,DUOX1,2)产生活性氧,以前曾参与调节癌细胞的铁死亡。NOX2在中性粒细胞中高度表达。作者假设NOX2是ROS的来源,在补充AA + DHA的中性粒细胞中引发脂质过氧化和铁死亡。通过进一步研究证明,减少PUFA-PL含量对于静止和活化的中性粒细胞来说都是必不可少的,以最大限度地减少由NOX2依赖性ROS产生驱动的脂质过氧化和铁死亡。

图4. T细胞和中性粒细胞中的脂质补充改变了对脂质过氧化和铁死亡的敏感性
(图源:Morgan, Pooranee K et al., Nat Cell Biol. 2024)
三、小结
作者构建了人类和小鼠免疫细胞脂质图谱,检测到超过500种脂质,它们在宿主免疫中具有不同的作用,表明免疫细胞的脂质组成是不同的。此外,细胞成熟和活化改变免疫细胞脂质组成,而关键的脂质特征是在免疫细胞发育过程中获得的。通过分析定义脂质特性和功能的主要化学特征(例如PL头群、酰基链多样性、化学连接),作者已经确定了独特的、免疫细胞特异性的脂质谱,并提供证据表明这些差异具有重要的生物学功能。具体来说,通过脂质图谱,作者发现PUFA-PLs丰度的差异为之前描述的T细胞对铁死亡的易感性以及骨髓细胞(包括中性粒细胞和单核细胞)对铁死亡的抗性提供了潜在的分子基础。
拜谱小结
许多特征定义了免疫细胞的身份,从物理参数,如细胞大小、核形态和颗粒含量,到分子特征,如转录组和蛋白质表面标记的表达。重要的是,这种差异是不同免疫细胞类型独特功能特性的基础。实际上免疫系统细胞的脂质景观也是免疫细胞身份的一个决定性特征,并且脂质组成的差异有助于细胞特异性功能。该研究通过基于质谱的靶向脂质组学技术构建了一个人类和小鼠免疫系统的综合“脂质图谱”,并揭示脂质图谱是免疫细胞身份的关键特征以及细胞特异性脂质表型是免疫细胞生理功能的基础。广泛靶向代谢组学是一种整合了非靶向代谢组学“广泛性”和靶向代谢组学“准确性”优点的新型代谢组学检测技术,具有高通量、高灵敏度、广覆盖度、定性定量准确等特点。拜谱生物已经推出了MLT4500医学高通量靶向脂质组产品,多针检测4500+脂质分子,其中甘油磷脂类3100+,提供绝对定量数据和丰富的数据分析内容,适用于人、大鼠、小鼠等动物来源的样本,包括血液、细胞、组织等样本类型。敬请咨询!
参考文献:Morgan PK,Pernes G,Huynh K, et al. A lipid atlas of human and mouse immune cells provides insights into ferroptosis susceptibility. Nat Cell Biol. 2024;26 (4):645-659. doi:10.1038/s41556-024-01377-z





关注公众号





关注小红书


