

400-820-8531




肿瘤放化疗抵抗是临床肿瘤治疗的一大棘手难题,而DNA损伤应答的异常激活,正是肿瘤细胞逃脱放化疗杀伤、导致治疗失效的核心原因。作为调控蛋白功能的重要翻译后修饰,棕榈酰化在DNA损伤应答中的具体调控机制,此前一直未被完全阐明。
近日,重庆大学附属肿瘤医院徐波团队在Oncogene期刊上发表了题为“ZDHHC20-mediated S-palmitoylation of KAP1/TRIM28 promotes DNA damage repair”的研究文章。首次解析出ATM-ZDHHC20-KAP1轴调控DNA损伤应答的分子机制,证实ZDHHC20可棕榈酰化修饰KAP1的C232位点,ATM通过磷酸化ZDHHC20的S339位点激活其酶活性。为肿瘤放化疗增敏找到了全新潜在靶点。拜谱生物为该研究提供了棕榈酰化修饰蛋白质组学技术服务。

研究结果
1、ZDHHC20参与DNA损伤反应(DDR)
为了阐明DNA损伤反应(DDR)中棕榈酰化的调控机制,对23种ZDHHC蛋白进行了敲除/过表达(图1A),CCK-8、集落形成、WB检测等实验证明ZDHHC20在DDR中的作用最大(图1B-1I)。免疫荧光实验显示,双链断裂标志物γH2AX焦点数量显著增加(图1J-1K),说明未修复双链断裂数量显著更多,DNA损伤修复过程延迟。这些结果表明,ZDHHC20可能在DNA损伤应答中发挥潜在的调控作用。
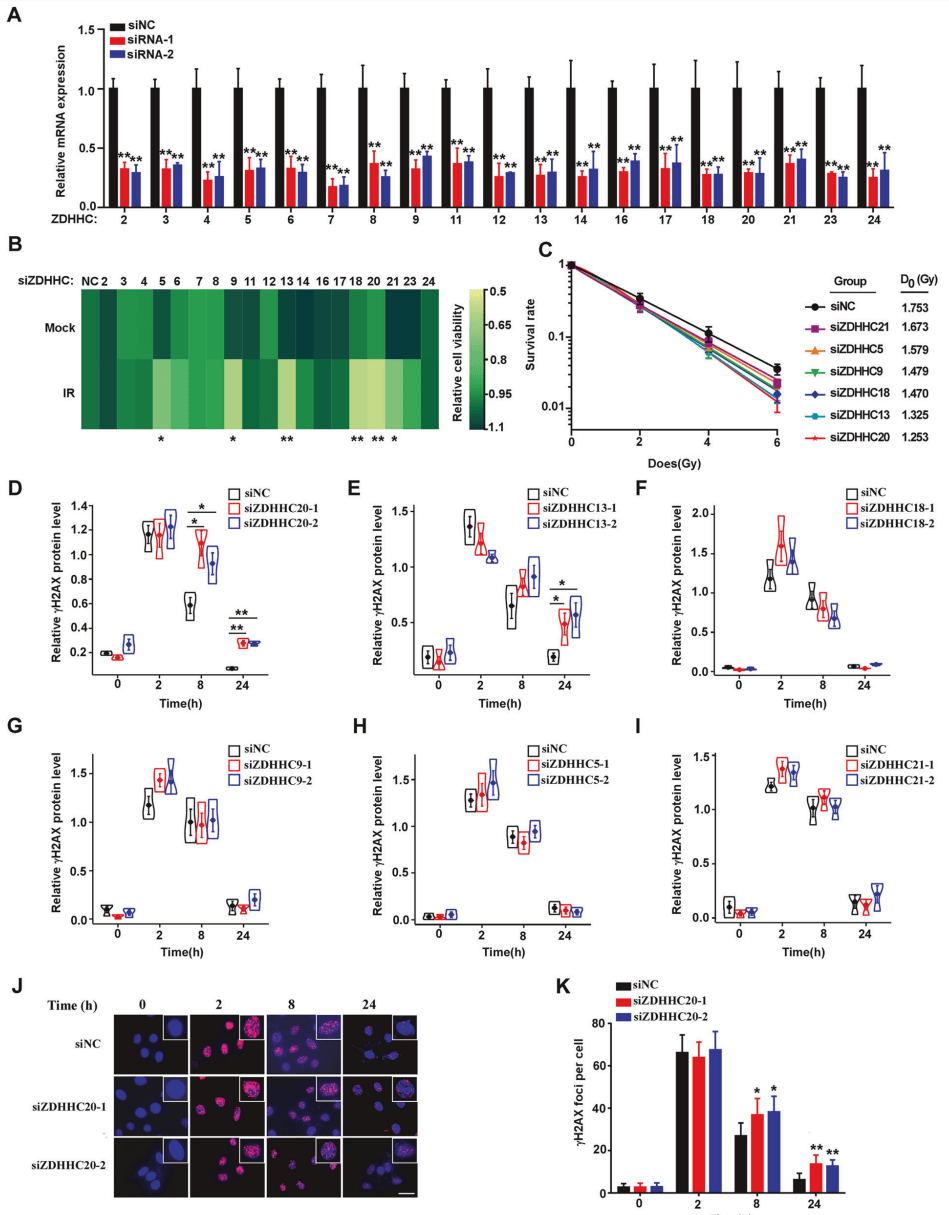
图1 ZDHHC20是主要参与DDR的关键棕榈酰转移酶之一
2、ZDHHC20调控对放射的敏感性和DNA损伤修复
为了确认ZDHHC20的作用机制,构建Zdhhc20敲除鼠模型,并进行全身IR处理,发现敲除鼠的存活时间显著低于正常鼠(图2A-2C)。测量对IR敏感的小肠上皮,发现敲除鼠的小肠隐窝明显缩短(图2D)。体外构建ZDHHC20敲除细胞系(HeLa和U2OS),发现其存活率显著降低(图2E-2F)。裸鼠异种移植模型显示,在IR处理下,ZDHHC20敲除的HeLa细胞生长速率显著降低(图2G-2H),肿瘤重量显著减少(图2I),γH2AX增加(图2J-2L),彗星尾显著延长(图2M-2N)。利用EJ5和DR的U2OS细胞系,发现敲低ZDHHC20会显著减弱这两种细胞系的修复能力,表明ZDHHC20可能通过干扰NHEJ和HR影响DNA修复(图2O)。由此说明,抑制ZDHHC20的表达可在体外和体内提高放射敏感性,同时抑制DNA损伤的修复。

图2 ZDHHC20调控对放射的敏感性和DNA损伤修复
3、ZDHHC20促进染色质重塑因子KAP1的棕榈酰化
为了确定ZDHHC20的下游靶蛋白,对WT和ZDHHC20敲除的HeLa细胞进行棕榈酰化修饰蛋白组检测,结合Co-IP分析,确定6种可能受ZDHHC20调控的下游靶蛋白。通过文献调研和Co-IP证实,ZDHHC20与染色质重塑因子KAP1存在互作,且会因离子辐射(IR)而增强(图3B-3F)。免疫荧光显示,ZDHHC20蛋白与KAP1在细胞核中共定位(图3G)。
棕榈酰化蛋白质组表明,ZDHHC20敲除后KAP1的棕榈酰化水平显著降低(图3H)。ABE有类似的结果,KAP1可发生棕榈酰化且受到IR的进一步诱导(图3I),ZDHHC20敲除后KAP1的棕榈酰化水平降低(图3J)。位点分析显示,KAP1的232位半胱氨酸可发生棕榈酰化(图3K),点突变后发现其棕榈酰化水平显著降低(图3L)。
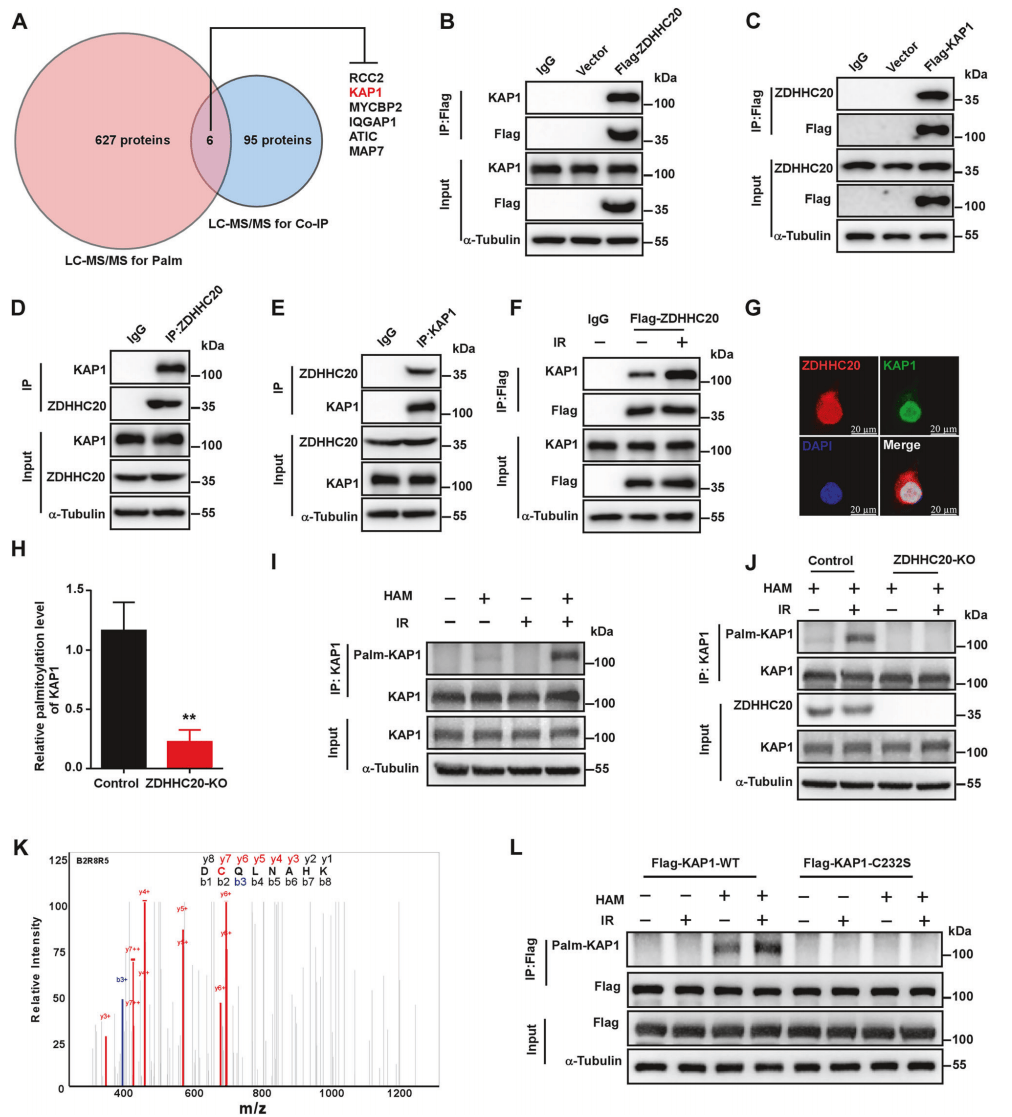
图3 ZDHHC20促进KAP1的棕榈酰化
4、ZDHHC20对DDR中KAP1的功能至关重要
基于先前研究结果,推测棕榈酰化可能会影响磷酸化KAP1与染色质的结合。细胞分级分离实验证明,ZDHHC20敲除和C232S点突变细胞中的p-KAP1在染色质上的定位减少(图4A-4B)。为了进一步证实影响KAP1与染色质的结合,利用ChIP-qPCR发现,C232S点突变细胞中KAP1在靶基因启动子的富集显著减少(图4C-4D)。敲除和点突变都显著抑制IR诱导的染色质松弛标志物Sat2和α-sat的表达(图4E-4H),染色质可及性降低(图4I-4J)。ATAC-seq分析表明,DDR期间C232S点突变细胞的开放染色质区域显著减少(图4K)。由此表明,ZDHHC20介导的KAP2棕榈酰化对其染色质重塑功能至关重要
此外,ZDHHC20敲除细胞中DNA损伤修复蛋白BRCA1和53BP1向DNA损伤位点的募集显著减少(图4L-4M),在染色质上的定位减少(图4N)。由此说明,ZDHHC20对于KAP1在DNA损伤反应(DDR)中的作用是不可或缺的。
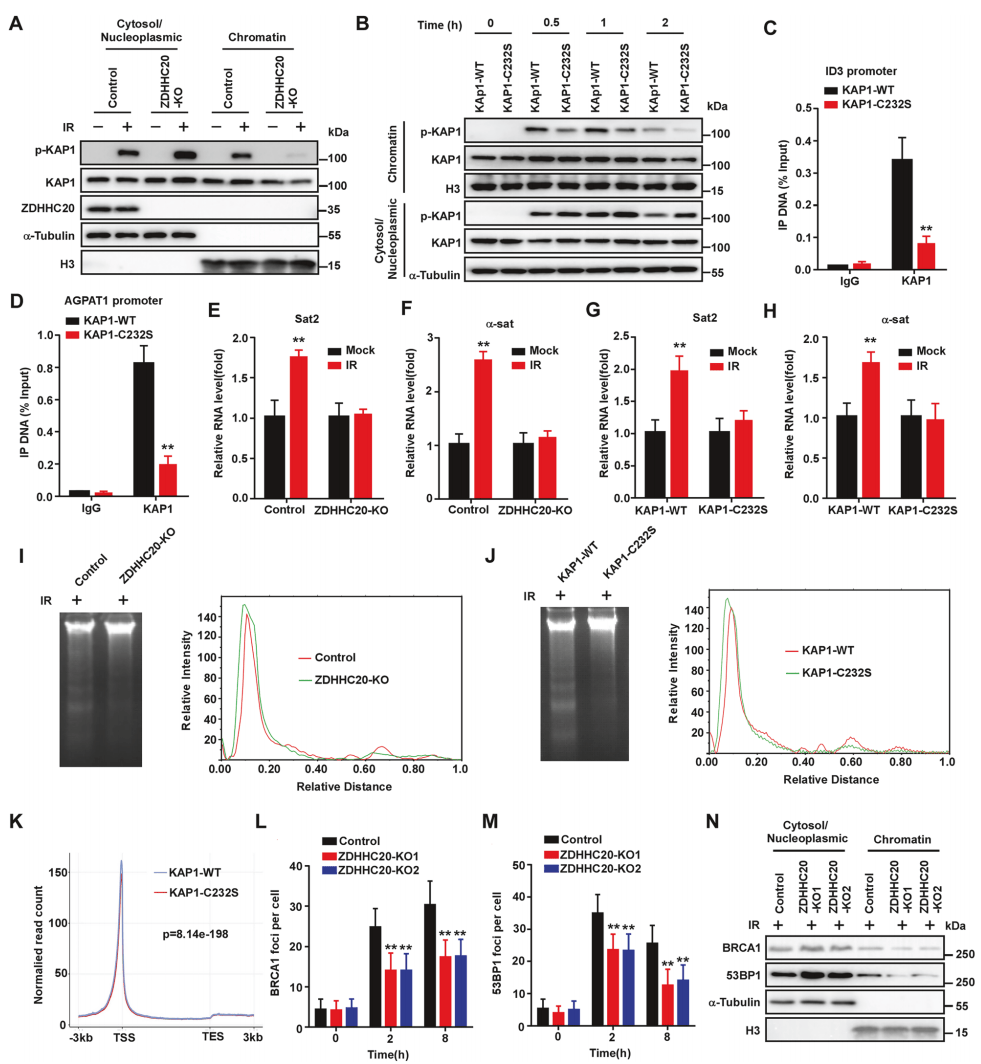
图4 ZDHHC20的缺失削弱了KAP1在DDR中的调控作用
5、ZDHHC20介导的KAP1 C232位的棕榈酰化是DNA损伤修复所必需的
相比较正常细胞,C232S点突变细胞的DNA损伤修复能力显著下降(图5A-5E)。同时敲除ZDHHC20则可消除两组细胞在DNA修复能力上的差异(图5F-5H)。此外, C232S点突变细胞表现出更高的辐射敏感性(图5I)。裸鼠异种抑制模型显示,C232S点突变的肿瘤生长速度显著减慢,肿瘤重量明显减轻(图5J-5L)。
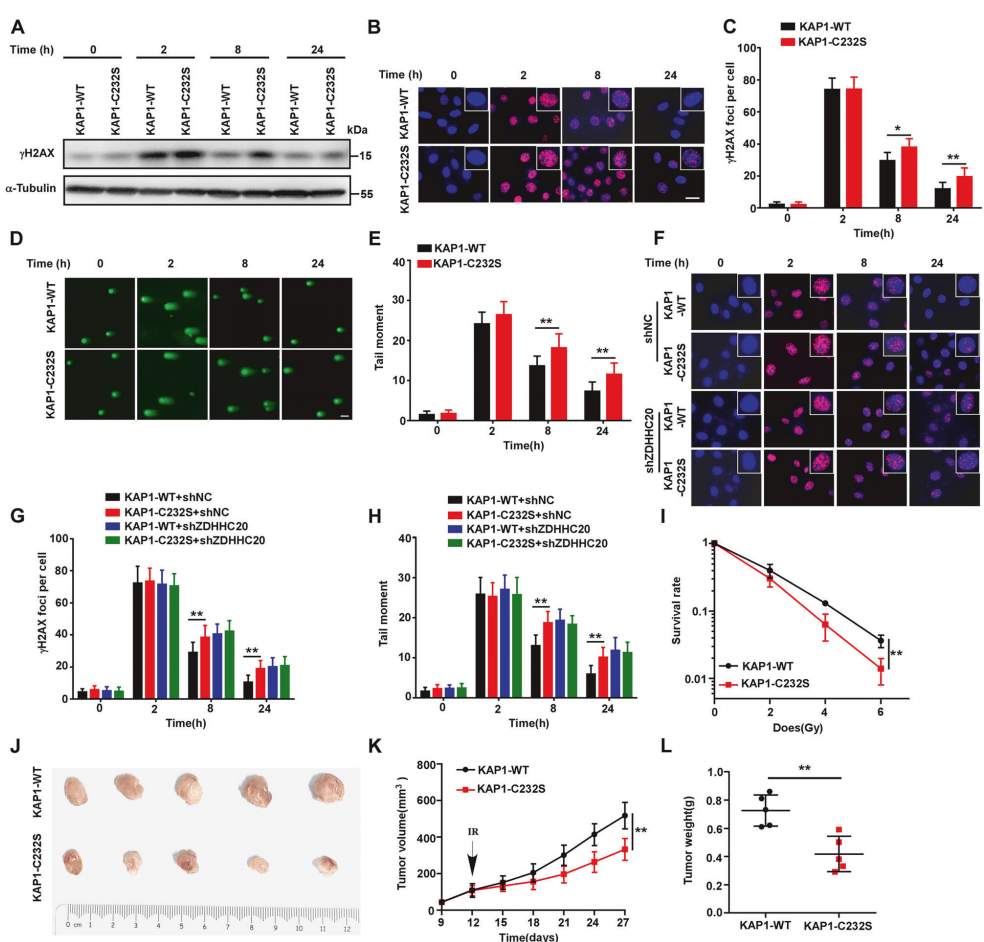
图5 KAP1 C232S导致肿瘤细胞的DNA损伤修复缺陷
6、ZDHHC20介导的KAP1棕榈酰化依赖于ATM
有研究证明ZDHHC20是激酶ATM的磷酸化底物,且339位的丝氨酸可能是被ATM磷酸化的位点,需要进一步探究ATM对ZDHHC20介导的棕榈酰化的调控作用。首先,通过Co-IP证明ZDHHC20可与ATM结合(图6A-6C),磷酸化特异性抗体检测证明ZDHHC20存在磷酸化,且IR处理后,磷酸化水平显著升高(图6D)。
点突变ZDHHC20 S339A,发现其磷酸化水平显著降低(图6E),且抑制其与KAP1的互作(图6K),减弱KAP1的棕榈酰化(图6M)。敲低ATM或通过Ku55933抑制ATM活性,都能显著抑制ZDHHC20的磷酸化(图6F-6G),并减弱其与KAP1的互作(图6I-J),抑制KAP1的棕榈酰化(图6L),p-KAP1与染色质的结合显著减少(图6N)。ATM可特异性地识别ZDHHC20的SQ基序,该基序在多个物种中具有进化保守性(图6H)。由此说明,DDR过程中ZDHHC20介导的KAP1棕榈酰化依赖于ATM。
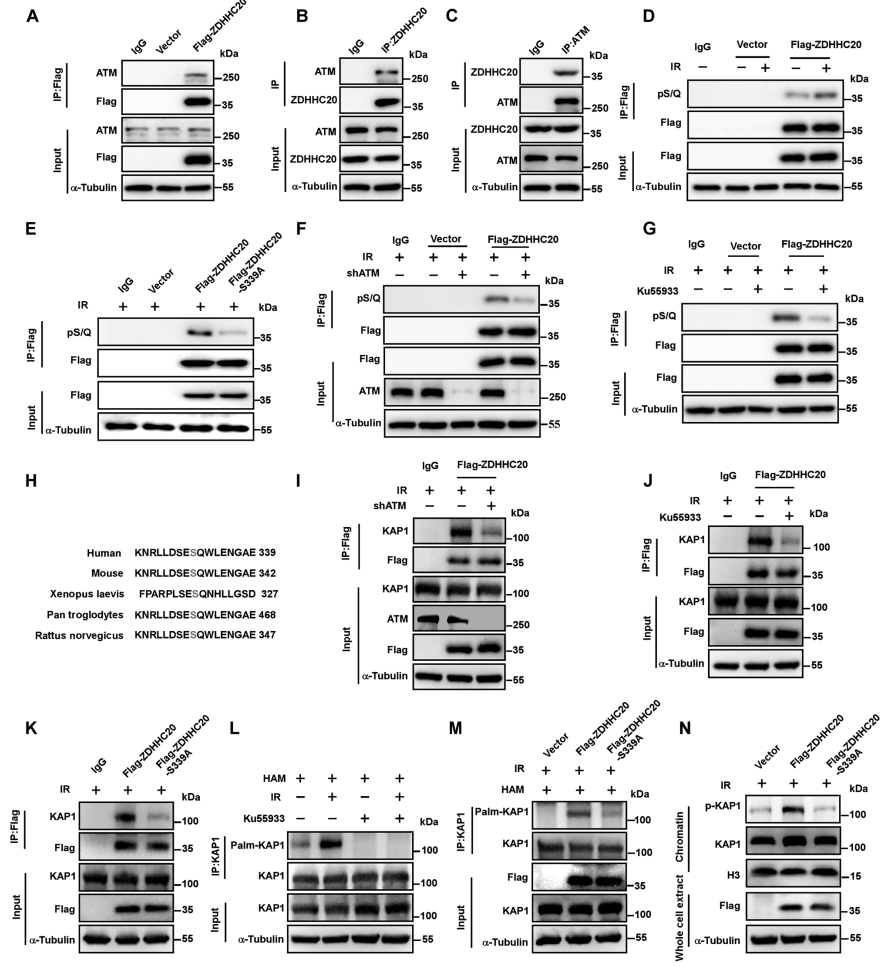
图6 ZDHHC20介导的KAP1棕榈酰化依赖于ATM
7、ZDHHC20在S339位点的磷酸化是DNA损伤修复所必需的
为了进一步确认ZDHHC20的 S339位点的磷酸化在DDR中的作用,将连有flag的WT和点突变S339A ZDHHC20回补到ZDHHC20敲除的HeLa细胞中(图7A),发现点突变细胞的DNA损伤修复能力显著降低(图7B-7F)。将点突变S339A ZDHHC20回补到U2OS的两种细胞系,发现点突变细胞的非同源末端连接(NHEJ)和同源重组(HR)的修复活性均降低(图7G-7H)。此外,点突变细胞对放射的敏感性增加(图7I)。由此表明,S339位点的ZDHHC20磷酸化在DDR中起着关键作用。
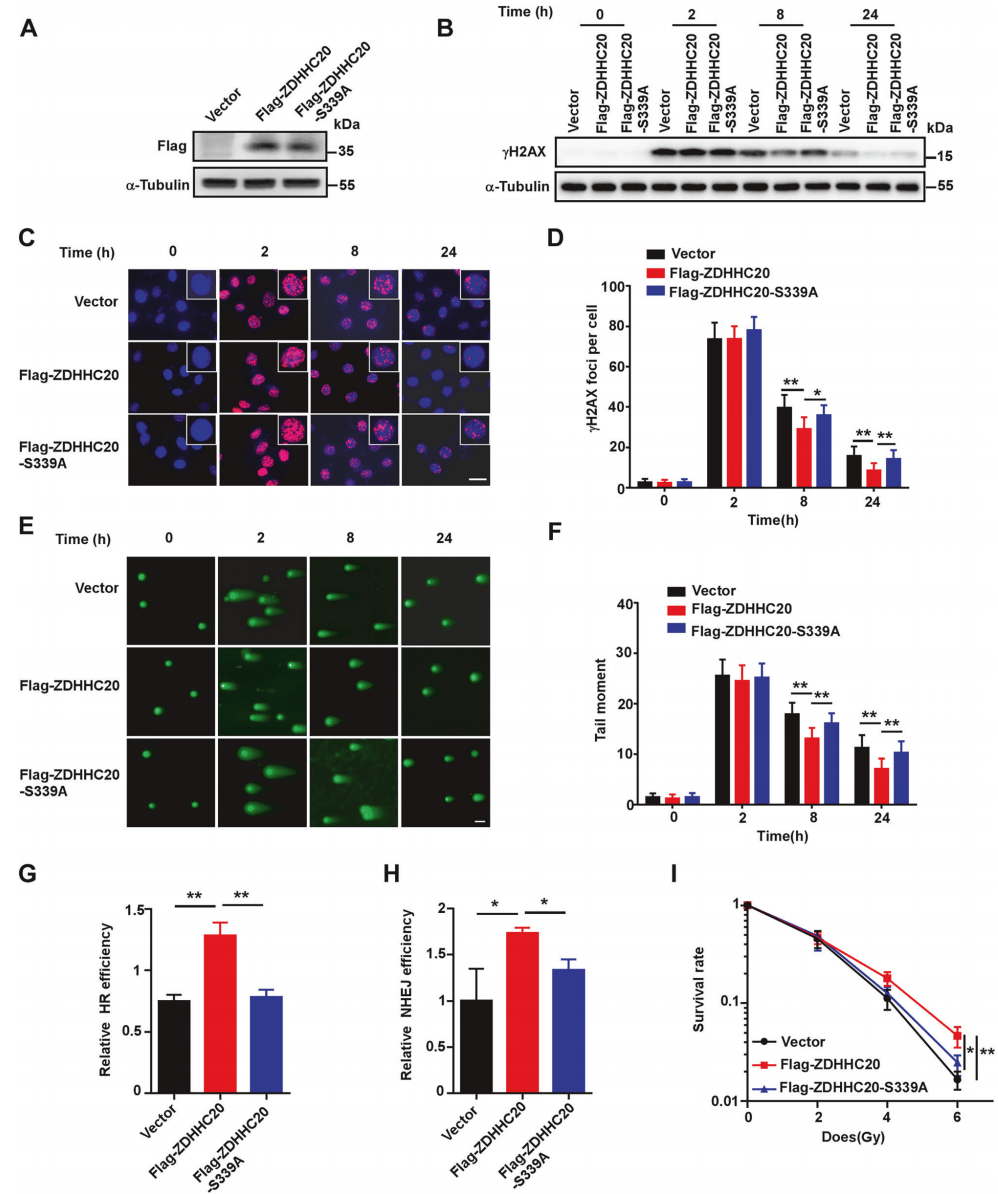
图7 ZDHHC20 S339A突变抑制DNA损伤修复并增加肿瘤细胞的放射敏感性
文章小结
本研究通过细胞实验、基因敲除小鼠及异种移植瘤模型,结合棕榈酰化修饰蛋白质组学等技术证实,ZDHHC20 是调控 DNA 损伤应答的核心棕榈酰基转移酶。就下游机制而言,KAP1是ZDHHC20的调控底物,在其 C232 位点进行棕榈酰化修饰,并增强磷酸化 KAP1 与染色质的结合。上游调控信号探究发现,DNA损伤诱导ATM激酶在ZDHHC20的S339位点发生磷酸化修饰,该修饰能提高 KAP1 的棕榈酰化水平(图8)。
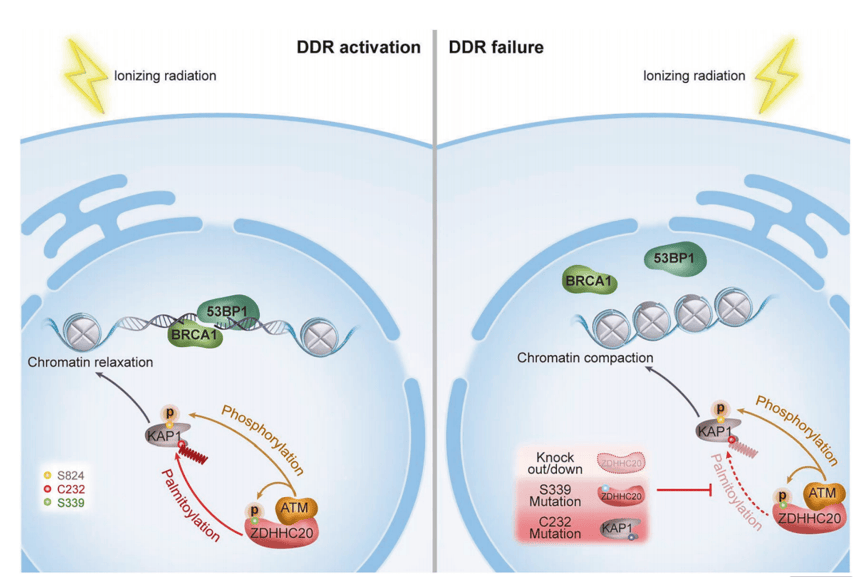
图8 作用机制图
拜谱小结
棕榈酰化是重要的脂质化修饰形式,一般发生在半胱氨酸残基上。棕榈酰化修饰最重要的功能是增强可通行蛋白与膜的亲和性,从而调控蛋白质的定位与功能。
拜谱作为国内领先的多组学服务公司,可提供基因组、转录组、蛋白组、翻译后修饰组、代谢组等多组学服务。围绕半胱氨酸氧化还原修饰,拜谱生物可提供产品全面、技术体系成熟的氧化还原修饰系列产品,包括棕榈酰化、亚硝基化、谷胱甘肽化、硫巯基化、次磺酸化、total氧化还原、游离巯基,助力一系列文章发表,积累了丰富的项目经验,欢迎大家咨询,获取专属棕榈酰化研究方案与技术资料!
参考文献
Liu Z, Wang H, Han Y, Chen Y, Wang Y, Zhang S, Mo Y, Wang C, Xiao M, Xu B. ZDHHC20-mediated S-palmitoylation of KAP1/TRIM28 promotes DNA damage repair. Oncogene. 2025 Dec;44(46):4533-4548. doi: 10.1038/s41388-025-03604-9. Epub 2025 Oct 18.





关注公众号





关注小红书


